
Evolution has given us many unusual animals. Some of them are so unusual that their discoverers were called crooks (as was the case with the platypuses in 1797). But not only the fauna can boast of non-standard species, among the representatives of the flora there are also white crows, namely, plants that do not want to sit on a sunny diet, but prefer juicy insects and other small animals as a dish of the day. Earlier, we already got acquainted with a study in which scientists examined the Venus flytrap and its marginal thorns.... Today we have to find out how sensitive these thorns are. Scientists from the University of Zurich (Switzerland) conducted a series of practical experiments, the purpose of which was to measure the speed of the venus flytrap's reaction to the touch of a particular force. How exactly did the scientists tickle the flycatcher, how quickly did it react, and how do these results explain the predatory plant's gastronomic preferences? We will find answers to these questions in the report of scientists. Go.
Research basis
In order not to stretch this narrative, the description of the Venus flytrap (which was already used in the previous article) is hidden under the spoiler:
What is a flycatcher?
, . , . — , . , «» (, ) ( , ). 630 , , .
: . ( ), — , .

. , — ( 300 000 1 ) , . , , . , . , .
, «».
, . : , , . , , . , , , . -, . .

: , - .
, -, . . , . , . , . , ? , . 2 20 . , , . , , , , .
. , . , , . , ( ), , . . , , . — , . , . , 2-3 .
. , , . , , . , .
True Facts: Carnivorous Plants ( : ). .
, -. , , .
, , , , « ». , , «». , , , , . , . , .
: . ( ), — , .
. , — ( 300 000 1 ) , . , , . , . , .
, «».
, . : , , . , , . , , , . -, . .
: , - .
, -, . . , . , . , . , ? , . 2 20 . , , . , , , , .
. , . , , . , ( ), , . . , , . — , . , . , 2-3 .
. , , . , , . , .
True Facts: Carnivorous Plants ( : ). .
, -. , , .
, , , , « ». , , «». , , , , . , . , .
The main idea of the mechanism of triggering the trap of the venus flytrap is that the future prey needs to make two touches to the sensitive hairs of the plant within 30 seconds. Each touch of them produces an action potential * required to close the trap. Scientists have believed this for 200 years, but is the theory consistent with practice?
Over the years of research, scientists have deduced a scenario of events that should (in theory) unfold during a flycatcher's hunt:
- a hungry plant attracts insects by secreting volatile compounds;
- not very savvy insect explores the plant (hoping for free food), during which it touches one of the six sensitive hairs, thereby causing an action potential (AP from action potential);
- 30 , «It’s a trap!», , ;
- (, ), (C12H18O3) .
As a result, the previously opened "petals" of the flycatcher turn into a kind of stomach, where unsuccessful prey will be digested.
In this work, scientists decided to consider in detail the process of converting mechanical stimulation of sensitive hairs into an electrical signal. We know for sure that touching sensitive hairs opens mechanosensitive ion channels, but we have not yet been able to identify these channels.
While these putative channels are open, the receptor potential * (RP from receptor potential ) increases, and if the deflection of the hair is large enough, RP reaches a threshold above which AP is manifested.
* — , .
* — , .
* — .
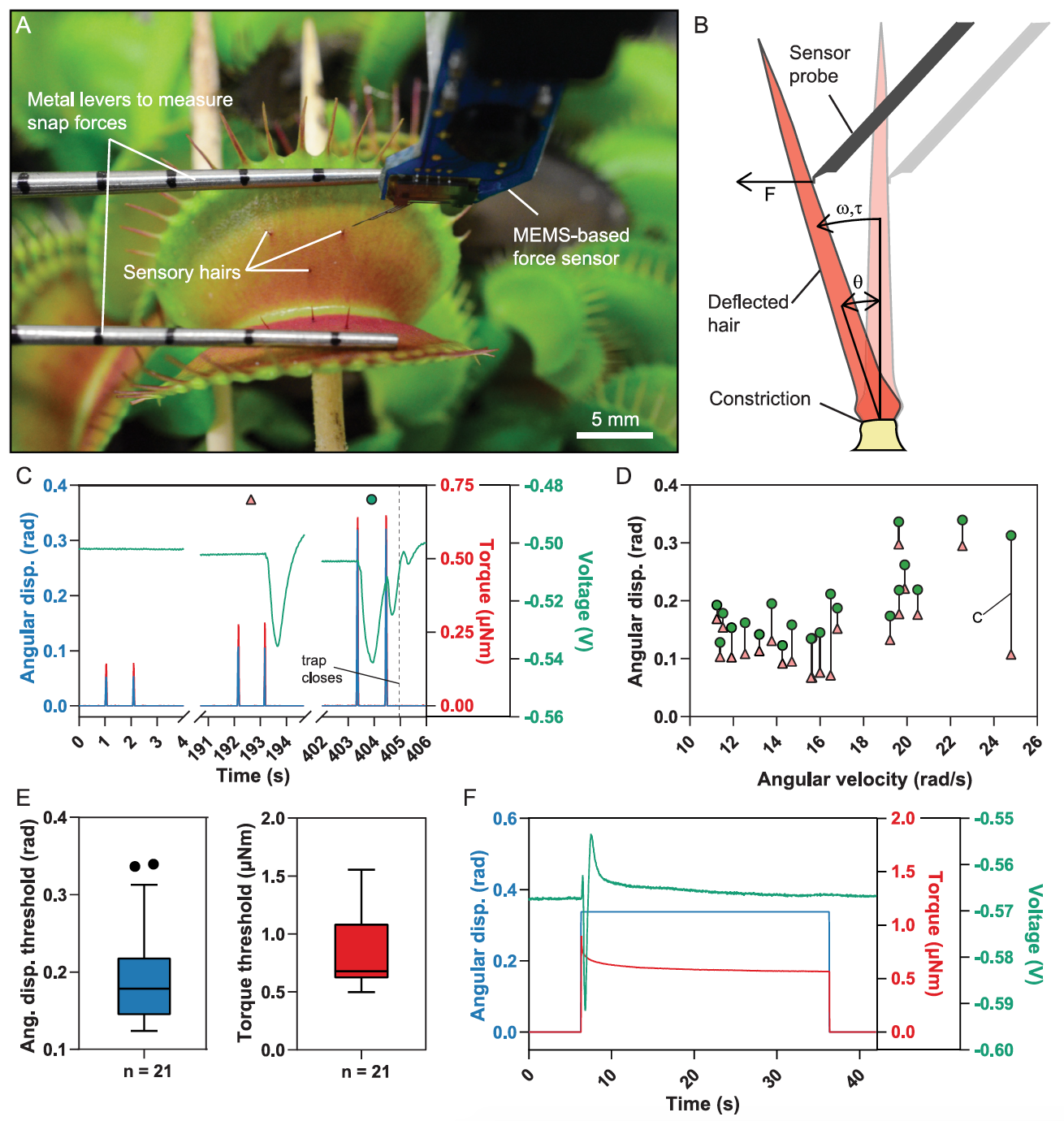
Image # 1
There have been earlier attempts to link mechanical stimuli and action potential generation, but there were no suitable instruments for accurate measurements. In the case of the study we are reviewing today, the scientists used a MEMS * -based force sensor mounted on a microrobotic system to precisely control the speed and amplitude of deflection, and simultaneously measure the applied force ( 1A and 1B ).
MEMS * (microelectromechanical systems) are devices that combine microelectronic and micromechanical components.
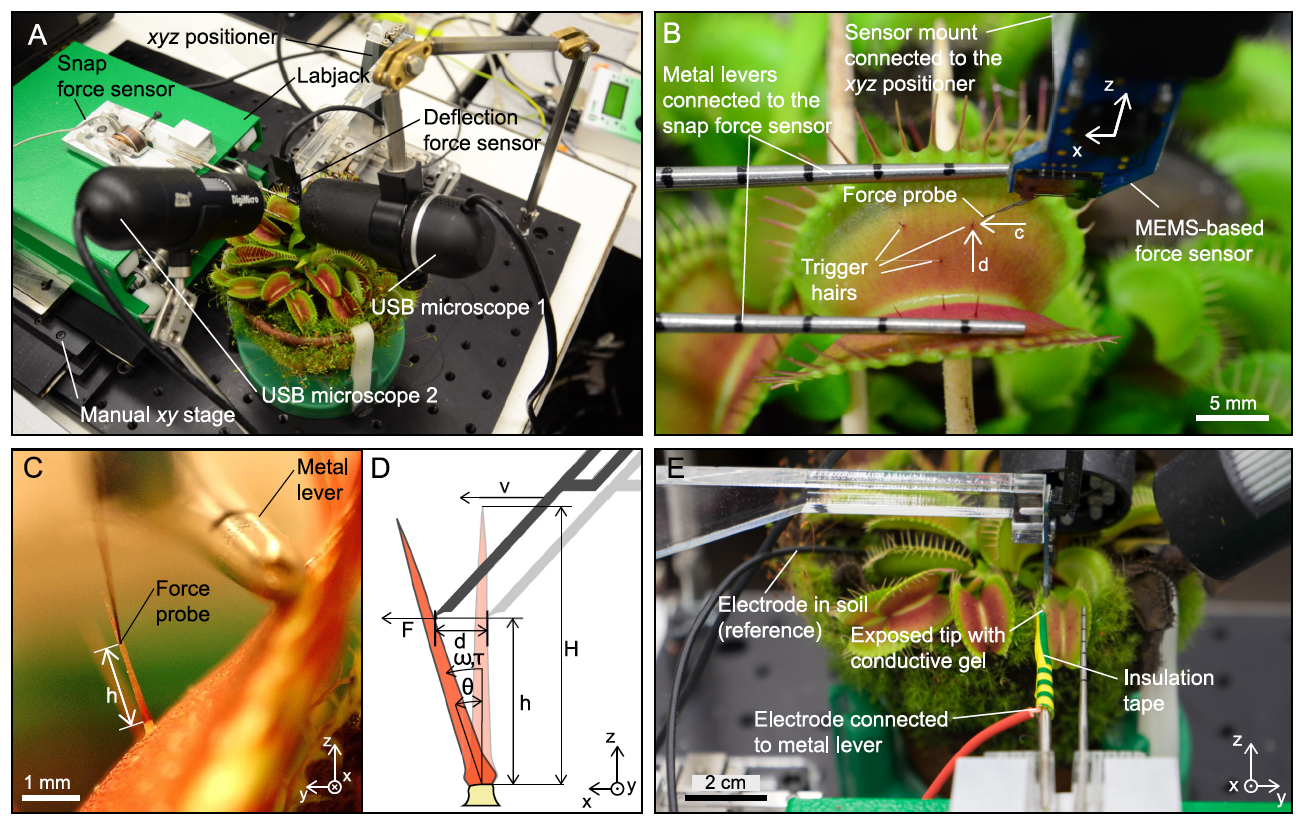
Image # 2
In this way, the scientists were able to accurately determine the range of parameters in which the deflection of the hairs leads to the closure of the trap, while the second force sensor measured the generated instantaneous force (image above).
Research results
During the experiments, the scientists took into account that spiders, flies and ants (common prey for a flycatcher) will cause sensitive hairs to deflect rather quickly. Consequently, the microrobotic system was deployed at full speed to mimic these stimuli. This resulted in a high initial angular velocity in the range of 10 to 20 rad / s.
Previous studies have found that ants deflect sensitive hair at an angular velocity of 0.25 - 7.8 rad / s, but slower than flies. Therefore, the parameters used are higher than the real ones, which is well suited for assessing the capabilities of the Venus flytrap.
At such high velocities, the duration of deflection is significantly shorter than other time-dependent factors involved, such as RP decay and sensory hair relaxation (1F ).
Taking into account the angular rather than linear deviation made it possible to correct the differences in the height of the contact of the sensory probe relative to the place of narrowing of the sensory hair, as well as the differences regarding different geometries of the sensory hair ( 1B , 2C and 2D ). Hence, one deflection can be approximated by a discrete increase in angular offset, and AP triggering is primarily dependent on the magnitude of the angular offset.
One deflection was defined as a combination of forward-backward angular displacement, similar to what occurs when a prey touches a hair. Each measurement consisted of two subsequent deflections with an interval of 1 s between them up to a given angular displacement θ. If the trap did not close, a wait phase (2 minutes) followed to ensure that the RP was completely reset. The wait for exactly two minutes, and not more / less, is due to the fact that a series of earlier experiments demonstrated that at temperatures below 30 ° C, two deflections of the hair within 30–40 s are required to quickly and completely close the trap.
After the waiting stage, the procedure was repeated with increasing angular displacements until the trap closing mechanism was triggered ( 1C and 1D). This occurred when the average displacement threshold θ = 0.18 rad or the average torque threshold τ = 0.8 μN m (n = 21) was reached.
Scientists note that the closure of the trap was never recorded if θ was below 0.12 rad and τ was below 0.50 μN · m ( 1E ). Consequently, these indicators are the lower limit of the angular deviation that is required to trigger the closure of the trap under the conditions of this experiment. It was also possible to determine that for the trap to operate, the insect must apply a force (F) of 0.5 mN near the tips of the hairs or 5 mN near the base.
AP measurements provided a link between sensory hair deflection and trap closure. When two consecutive deviations were well below the bias threshold (θ << 0.12 rad), no AP was observed. For deflection amplitudes near the displacement threshold (θ <0.12 rad), a single AP was detected after the second deflection.
This indicated that both hair deflections contributed to RP and that the AP induction threshold was only reached at the second deflection. As expected, a single AP was not enough to trigger the trap.
The assumption that each touch triggers AP is only valid if the deflection of the sensory hairs exceeds the displacement threshold. If so, then two APs (one for each hair deflection) were generated and caused the trap to close ( 1C ).
Double deflection of sensitive hairs resulting in two APs and trap closure.
These results suggest that rapid deflection of sensitive hairs increases RP to a certain level that depends on the amplitude of angular deflection. RPs can stack and can summon AP after multiple deflections if they are below the deflection threshold. However, the generation of one AP per touch occurs only if the deflection of the sensitive hairs is above the deflection threshold.
In other words, the action potential will be generated only when the sensitive hair has been “disturbed” enough. But a one-shot action potential is not enough to trigger the closing of the trap. This fits well with the theory that flycatchers will not waste energy on closing the trap every time something or someone touches their hair. Otherwise, there may be many idle traps without any energy (i.e. nutritional) return (the prey managed to escape, the prey was too small, or it was not prey at all, but garbage).
Given that the increase in RP with multiple deviations is additive, it would be logical to assume that sustained (long-term) displacement of a hair can have a similar effect.
To test this hypothesis, the scientists rejected the flycatcher's sensitive hair beyond the angular displacement threshold and locked it in that position for 30 seconds ( 1F ). However, this did not trigger the traps.
The initial displacement caused a single AP, after which the tension quickly returned to baseline, although the hairs remained deflected.
If the sustained bias was to contribute to RP, it would have to remain above the threshold, in which case an AP series was expected. Additional analyzes have shown that angular misalignment does play an important role in RP, but static hair deflection does not contribute in any way.
It is curious that in earlier experiments the situation was different: a single displacement of the hairs led to the closure of the trap. However, this was not an integral single displacement, but many small displacements, since the oscillations that are accompanied by manual deflection of a hair are probably greater than the angular displacement threshold.
The next stage of the study was to create an electromechanical charge buildup (ECB ) model , which surprisingly predicts the closure of traps from a single touch.
Based on the data obtained during the experiments, the scientists developed a simple model to study the limits of angular displacement and speed within which the traps will react.
In the ECB model, mechanical deflection results in charge accumulation RP as a function of angular velocity ω and displacement θ, while charges are continuously dissipated. If the accumulated charges exceed a certain threshold Q th RP , an AP occurs. Additionally, a refractory period t RP has been implemented , representing the time interval required for another AP to be activated.
As a result, the model predicted that if the deviations are too fast and / or too small, then more than two such deviations may be required to cause the trap to close (red area at 3A ).

Image # 3
This is due to the fact that one deviation is not enough to activate AP ( 3B), as shown by practical experiments ( 1C , center).
Similarly, the model showed that very low angular velocities (ω <0.04 rad / s) cannot fill RP.
But the most unexpected result of the simulation was the prediction of the range of intermediate angular velocities (0.04 rad / s <ω <10 rad / s), at which one deflection is enough to activate two or more APs, which are necessary to close the trap ( 3B ).
However, such a prediction contradicts the idea of how the venus flytrap trap mechanism works. Therefore, the scientists decided to check whether this model situation can be realized in practice.
A single deflection of sensitive hairs resulting in trap closure.
Surprisingly, trap closure did indeed occur with a single deflection of hairs if the angular velocities were low enough.
To narrow down the range of values in which this occurs, scientists repeatedly deflected the same sensory hairs at different angular velocities until the trap closed. There was a 2 minute gap between two successive deflections until the trap recovered and RP was depleted.
The lower limit of the angular velocity ω required to close the trap at one deflection was determined by gradually increasing the angular velocity after each deflection (n = 17). The initial velocity was below 0.009 rad / s, since the trap was never closed at this speed.
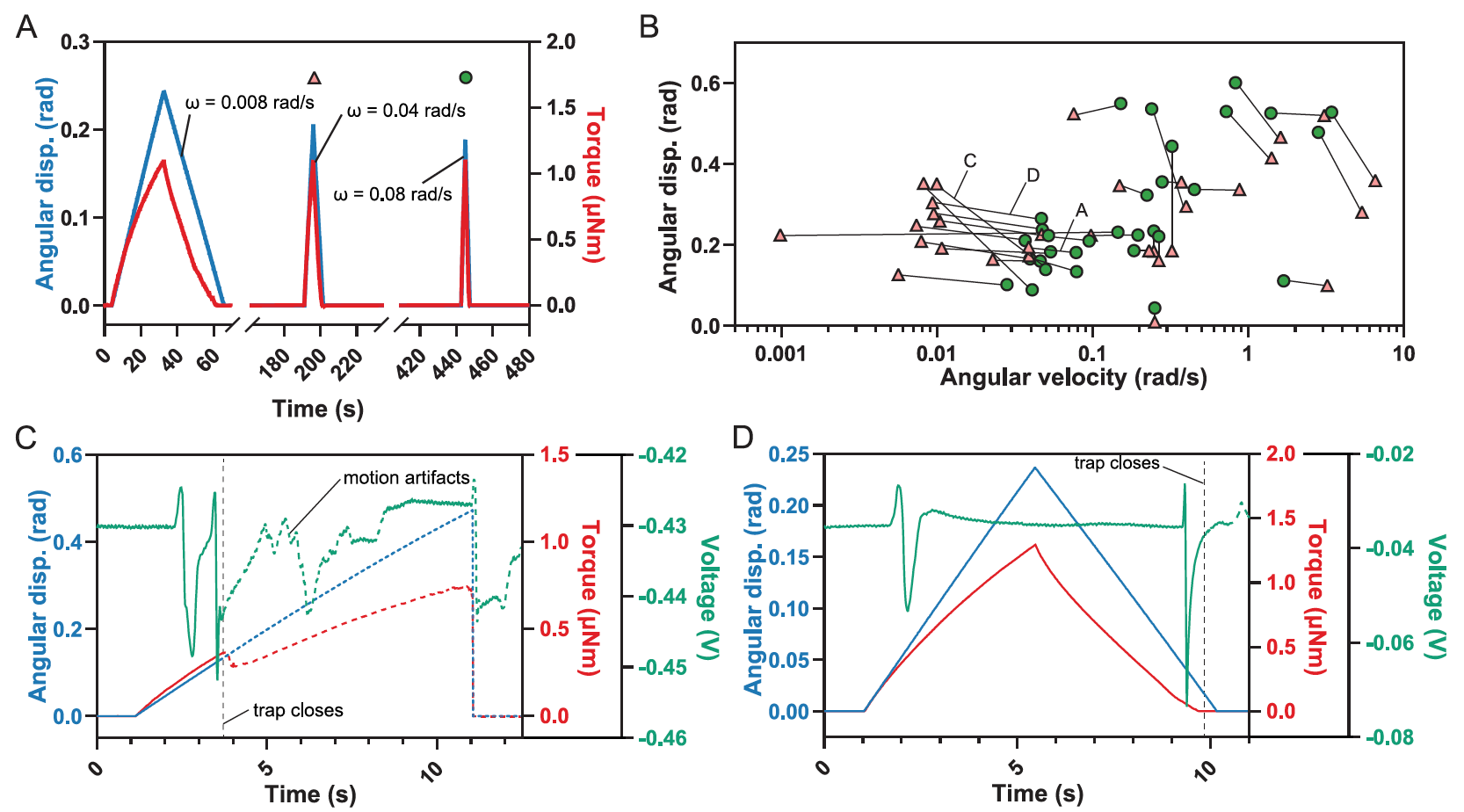
Image # 4
Subsequent single hair deflections were performed with a gradual increase in speed until the trap was triggered ( 4A ).
The upper limit was determined in a similar way, starting with a speed of ω> 3 rad / s followed by a stepwise decrease (n = 9).
In addition, another series of experiments with one deflection (n = 5) was performed, in which the velocity of the force probe was kept constant, which resulted in an intermediate angular velocity between 0.2 and 0.4 rad / s, while the angular displacement θ gradually increased during the time of subsequent deflections to obtain the lower bound θ required for the trap to operate under the conditions of a single deflection of hairs.
All single deflections leading to trap closure, together with prior stimuli for which no trap closure occurred, identified the region where a single deflection causes closure ( 4B ).
A single deflection can cause the trap to close at intermediate angular velocities of deflection (0.03 rad / s ≤ ω ≤ 4 rad / s). But deflection alone at lower or higher speeds will not be enough.
Experiments, where the rate of deflection of the hairs was rather low, showed that the trap is triggered both when the sensor approaches the base of the hair and when the sensor moves away from it.
Single deflection of sensitive hairs resulting in two APs during initial bending, resulting in trap closure.
When the trap was closed during the approach of the probe, two successive action potentials ( 4C ) were observed during the deflection of the hairs . When the trap was closed during the removal of the probe, one AP appeared at the moment of deflection of the hair, and the second after the removal of the probe, when the hair returned to its original position ( 4D ). In both cases, the second AR led to the immediate closure of the trap.
In conclusion, the scientists decided to measure the closing force of the trap ( 5A ).
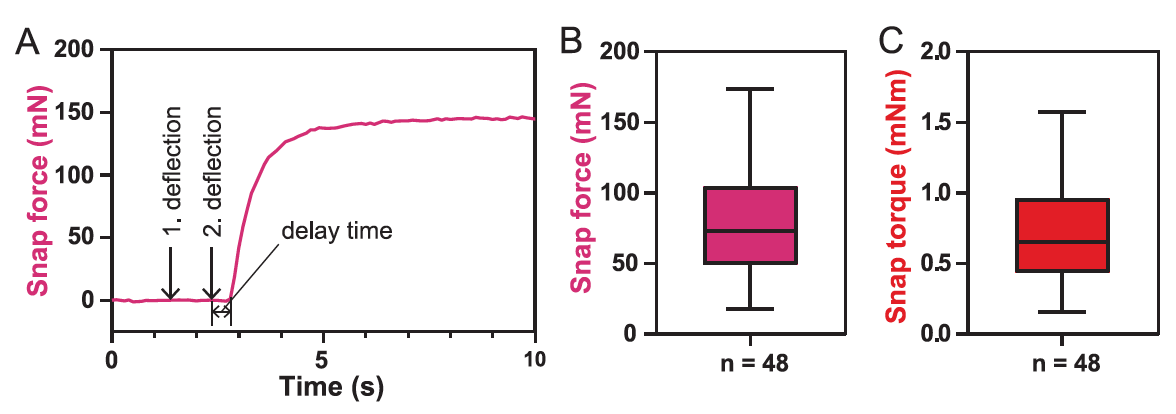
Image No. 5
For the 48 trap, the average value of the gripping force F close was 73 mN ( 5V), and the range of values was from 18 to 174 mN.
These values are lower than those previously published (140-150 mN). There is an explanation for this: in this case, the force was measured at the beginning of the closing of the trap, and in previous observations, the force acting on the edges of the leaf valves of the trap at the end of closing was measured.
Since the measured force is highly dependent on the position of the transducer and also on the orientation and size of the sheet, the closing torque τ close around the middle of the sheet with an average value of 0.65 mN · m is the best value for describing the trap closing force ( 5C ). The delay time, that is, the time between the mechanical stimulus and the beginning of the trap closing, was 0.6 ± 0.3 s.
For a more detailed acquaintance with the nuances of the study, I recommend that you look atreport of scientists .
Epilogue
The Venus flytrap does not run like a cheetah, does not have deadly poison like a cobra, and certainly does not hear its prey like an owl. But this plant is indeed one of the best carnivores on the planet, as its hunting mechanism works like a clock.
In this work, scientists were able to determine what these or those parameters should be for the trap to work. As it turned out, in most cases, to trigger the trap, the victim must make two successive mistakes: first, touch the flytrap's sensory hair; the second is to do it again.
The repeated deviation of the hair from its initial position leads to the generation of a second action potential, which starts the process of closing the trap. Nevertheless, simulation has shown that under certain conditions, one touch may be enough, but in reality this is most likely unlikely.
In other words, can the Venus flytrap serve as a mosquito hunter? No, because these bloodsuckers are very small and too nimble. But the flycatcher does not regret flies or snails.
Thanks for your attention, stay curious and have a great weekend, guys! :)
A bit of advertising
Thank you for staying with us. Do you like our articles? Want to see more interesting content? Support us by placing an order or recommending to friends, cloud VPS for developers from $ 4.99 , a unique analogue of entry-level servers that we have invented for you: The Whole Truth About VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps from $ 19 or how to divide the server correctly? (options available with RAID1 and RAID10, up to 24 cores and up to 40GB DDR4).
Is Dell R730xd 2x cheaper in Equinix Tier IV data center in Amsterdam? Only we have 2 x Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 TV from $ 199 in the Netherlands!Dell R420 - 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB - From $ 99! Read about How to build the infrastructure of bldg. class with the use of Dell R730xd E5-2650 v4 servers at a cost of 9000 euros for a penny?